


In the third installment in this series, Jonathan Birch looks at “horizontal transmission” and relatedness in bacteria.
In Part 1, we saw that cooperative behaviour is rife in the world of microorganisms. The fruiting bodies of myxobacteria and social amoebas are famous, but any bacterial biofilm is held together by vast amounts of discreet cooperation – the bacteria emit all kinds of molecular “public goods” to their neighbours.
This cooperative behaviour isn’t always altruistic, in the sense of implying a life-long cost to the reproductive prospects of the producer. It is often hard to tell whether the production of a public good is altruistic or not. But since these public goods are often energetically costly to produce, their production may well be altruistic in at least some cases.
So what stabilizes the production of these costly public goods against the threat of free-riding? The basic answer is probably a familiar one: relatedness. Many of a bacterium’s immediate neighbours are members of the same strain, and as such are clonally related to it. Although biofilms often contain multiple species, most of the cooperation happens within species – inter-species interactions are mostly competitive. So Hamilton’s ideas are as relevant here as they are to large, multicellular organisms.
Gene mobility
When we apply Hamilton’s ideas to bacteria, however, complications arise. The source of the trouble is the tendency of bacteria to exchange genes “horizontally” – for example, by transferring small packets of DNA called “plasmids” through a process of cell-to-cell contact called “conjugation”. A growing body of evidence points to the importance of this process to social evolution in bacteria. It turns out that many of the genes implicated in producing public goods are actually carried on plasmids.
What is going on here? The key point is that plasmid transfer is a replicative process: the plasmid is copied from one cell to another. So, after conjugating, two nearby organisms may share genetic material they did not share before. We can therefore see how, at least in principle, plasmid transfer can lead to genetic similarity between organisms.
Plasmid transfer is thus a source of “relatedness” in the broad sense that matters in social evolution theory. It’s a process that causes bearers of the genes for a social trait to cluster together, so that the effects of that trait fall differentially on other bearers of the relevant genes. Recently, experiments by Tatiana Dimitriu et al. have provided some experimental evidence that this really happens – plasmid transfer really does lead to higher relatedness, and really does allow a plasmid that encodes a public good to spread more rapidly than a plasmid that does not.
Why is that? Think about it this way: the altruism-encoding plasmid, by transferring horizontally, forcibly converts nearby free-riders into altruists. Because it targets nearby free-riders, not random free riders, it generates the sort of clustering together of altruists that allows altruism to become evolutionarily advantageous. When the public good is produced, the benefits fall differentially on other organisms who carry the genes for producing it.
This is easiest to imagine if we imagine the free-riders being forcibly converted into altruists early in the life-cycle – so that organisms who converted them have time to reap the benefits of having altruistic neighbours. But there’s also a more subtle (and harder to imagine) variant of the process in which the free-riders are converted late in the life-cycle.
In this variant, altruists first help their neighbours by producing a public good, and only later convert nearby free-riders, after they have already done their free-riding. This time, the benefits of altruism don’t fall differentially on current bearers of the genes for altruism, but they do fall differentially on potential future bearers of those genes – and that too might, in the right circumstances, be enough to make altruism evolutionarily advantageous.
Rethinking relatedness
This is already a rather subtle and difficult idea – but unfortunately the complications don’t end there. Gene mobility has foundational implications for social evolution theory. If we want a theory that fully incorporates the effects of horizontal gene transfer on relatedness, we need to change the way we think about relatedness. Here, in short, is why (see Chapter 6 of The Philosophy of Social Evolution for further detail).
In multicellular animals like humans or ants, relatedness can often be estimated fairly well by looking at genealogy – the extent to which interacting organisms have ancestors in common. This isn’t always the best way to measure relatedness, and in recent years genealogy-based measures have given way to statistical measures of genetic similarity estimated using molecular markers. But what is still taken for granted by all of these measures is that an organism’s genome remains stable over its life cycle, so that the time in the life cycle at which you measure relatedness doesn’t really matter – the same animal will always have the same genes.
When genes for social traits are being transferred horizontally, an organism’s genome is not stable over the course of its life-cycle, and the time at which you measure relatedness really does matter. In fact, what matters fundamentally is not whether the actor and recipient share genes at the time they interact, but whether the recipient is a potential future bearer of the actor’s genes – a potential candidate for transmitting those genes to future generations.
The easiest way to see this is to take a “plasmid’s eye view” on the process. If you’re a gene carried on a plasmid, you value your own host as a route to future generations – but you also value potential future hosts. It might still be in your evolutionary interests to help a nearby organism that is not yet carrying a copy of you, if it might, in the future, conjugate with your host and acquire a copy. The best measure of “relatedness” in this context is one that assigns value to potential future hosts not on the basis of their current genetic makeup, but on the basis of how likely they are to transmit a copy of you to their descendants.
This way of thinking about relatedness is fundamentally diachronic: relatedness is really about genetic similarity between one organism at the moment of gene expression and another organism at the moment of gene transmission, not about genetic similarity between two organisms at the same time. In other words, it is not a synchronic relation between two simultaneous “time-slices” of organisms, but a diachronic relation between two temporally separated time-slices. That is an unfamiliar, non-intuitive way of thinking about relatedness – but it is one we may have to get used to as we learn more about social evolution in bacteria.
In short, we tend to think that whether or not two organisms count as “genetically related” depends on their present (whether or not they share genes now) and on their past (whether or not they have parents or other ancestors in common). But what really matters in evolution is the future. To assess whether an altruistic behaviour will be favoured by selection, the fundamental question is not “Do the actor and recipient have recent ancestors in common?” or even “Does the actor genetically resemble the recipient now?” The question is: “Will this behaviour increase the representation in future populations of the genes it expresses?” In microbes, the answer can be “no” to the first two questions but “yes” to the third.
← Part 2Part 4 →
Jonathan Birch is Assistant Professor in the Department of Philosophy, Logic and Scientific Method, specializing in the philosophy of the biological sciences. Most of his work concerns the evolution of social behaviour. He is also interested in the evolution of morality, animal sentience, and the relation between sentience and welfare.
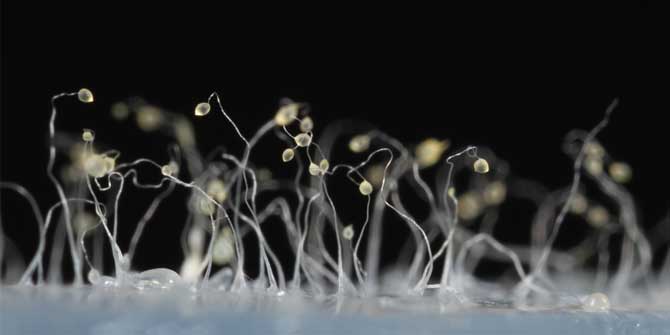
Featured image: Fruiting bodies of the myxobacteria, as sketched in 1892 by Roland Thaxter (compiled by the English Wikipedia). Public Domain.





































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}